Edit: this work has now been published as two papers. The first finds that mesoscopic beta-LFP oscillations may arise due to synchronization of rhythmic spiking in single neurons. The second explores how changes in synchronization relate to the diverse patterns illustrated in the poster below.
I'll present some of my ongoing thesis research at SfN as a poster this year. (This was originally titled "Identification of (~20 Hz) beta spatiotemporal dynamics in motor cortex LFPs".)
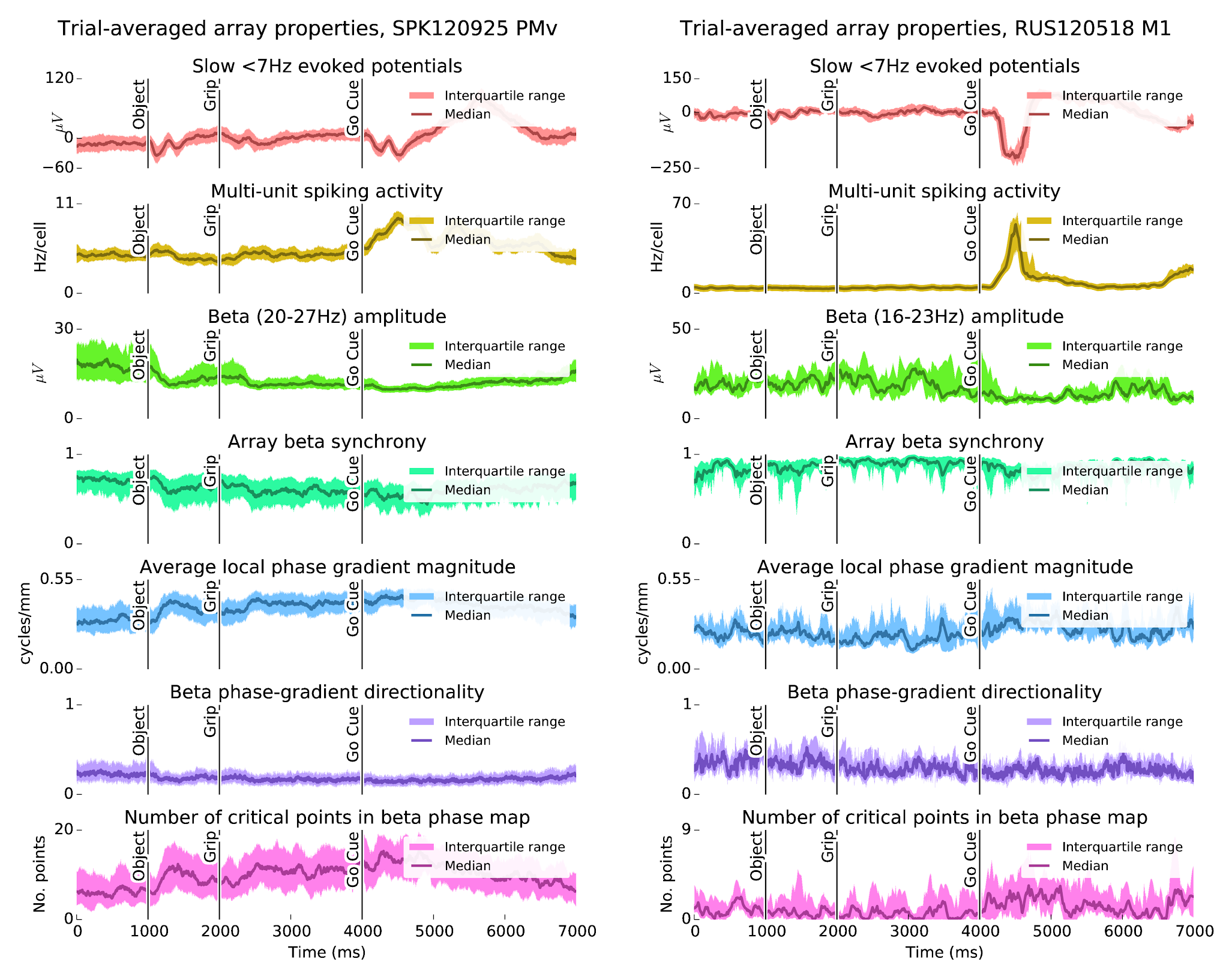
I've been looking into spatiotemporal waves in beta (~20 Hz) LFP oscillations during steady-state movement preparation. The wave dynamics seem to be more complex than previously reported [
1,
2]. Patterns vary depending on the properties of the beta-LFP oscillations, perhaps reflecting phase synchronization dynamics between local modules, or a signature of external input?
Abstract:
Modulation of beta (10-45Hz) oscillations is a prominent feature of primate motor cortex. Beta power is typically elevated during movement preparation, suppressed around movement onset, and enhanced during isometric force tasks. As shown by previous studies, beta oscillations can also appears as traveling waves in primate motor cortex. Understanding the mechanisms underlying the rapid modulation of beta LFP activity and the associated spatiotemporal patterns may shed light on the functional roles of these oscillations. It may also have important implications for movement disorders where regulation of motor cortex beta activity is abnormal (e.g. Parkinson's disease). Here, we examine motor cortex beta spatiotemporal LFP activity using multielectrode arrays (MEAs) in m. mulatta during a cued reaching and grasping task with instructed delay. Data from two monkeys are analyzed, each with a 96-MEA in ventral premotor cortex (PMv), and two 48-MEAs in the primary motor cortex (M1) and dorsal premotor cortex (PMd), respectively. Our main findings are threefold: (1) The transient nature of beta oscillation events together with variations in the beta band center frequency makes the identification of spatiotemporal structures challenging. In particular, different filtering and preprocessing steps can alter the apparent spatiotemporal dynamics. (2) Furthermore, attempts to summarize wave dynamics in terms of simple global structures, like plane waves, or rotating (radiating) waves around (from) a critical point, may fail to meaningfully describe the full range of beta spatiotemporal activity. (3) Despite these challenges, we find a variety of beta spatiotemporal patterns ranging from asynchronous states, i.e. states with no clear wave dynamics, to more locally synchronized states with complex wave dynamics, to globally coherent states. These globally coherent states may exhibit either traveling wave dynamics or homogeneous synchrony. We conjecture that the transitions among these different patterns may result from fast modulations of the effective lateral connectivity or from changes in spatiotemporal inputs to the cortical area.
This poster can be cited as
Rule, M. E., Vargas-Irwin, C., Donoghue, J., Truccolo, W. (2015) Identification of (~20 Hz) beta spatiotemporal dynamics in motor cortex LFPs. [Poster] Society for Neuroscience 2015, Oct 19th, Chicago, Il, USA.
Update: This work can now be found in the following papers
Rule, M.E., Vargas-Irwin, C.E., Donoghue, J.P. and Truccolo, W., 2017. Dissociation between sustained single-neuron spiking and transient β-LFP oscillations in primate motor cortex. Journal of neurophysiology, 117(4), pp.1524-1543.
Rule, M.E., Vargas-Irwin, C., Donoghue, J.P. and Truccolo, W., 2018. Phase reorganization leads to transient β-LFP spatial wave patterns in motor cortex during steady-state movement preparation. Journal of neurophysiology, 119(6), pp.2212-2228.