Update: Portions of these notes have now been published in the Journal of Neurophysiology as "Dissociation between sustained single-neuron spiking and transient β-LFP oscillations in primate motor cortex" and "Phase reorganization leads to transient β-LFP spatial wave patterns in motor cortex during steady-state movement preparation".
Task-locked modulations in neural activity
The Cued Grasp with Instructed Delay (CGID) task reliably elicits task-locked activity in all three motor areas (M1, PMd, PMv).
- Consistent with prior literature, the movement period of the CGID task is marked by slow motor evoked potentials (Fig. 2), increased single-unit firing rates (Fig. 3), and beta suppression (Fig. 4).
- Beta oscillations are enhanced during the first four seconds of the task, although there are some differences between subjects.
- The average level of beta-LFP synchrony is correlated with beta-LFP power, and varies across phases of the task.
- We find no evidence of task-locked phase resetting of beta LFP oscillations
- The spatiotemporal structure of beta-LFP waves is correlated with amplitude and synchrony, with lower amplitudes reflecting more complex wave structures, and higher amplitudes as more synchronous.
Figure 1: The CGID task reliably elicits evoked potentials, which correlate with beta suppression. In subject S, beta power is strongest in the first second before object presentation. In subject R, beta oscillations are more variable, with somewhat stronger power between the grip and go cues. In both animals, high beta power appears to correspond to periods of higher beta synchrony, and larger phase gradient directionality, a measure of how much LFP activity resembles a plane wave. Conversely, increases in the average magnitude of the Hilbert phase gradient, which summarizes how quickly beta phase changes over the array, and in the number of critical points in the Hilbert phase gradient, which summarizes the complexity of the beta spatiotemporal wave patterns, correspond to periods of beta suppression.
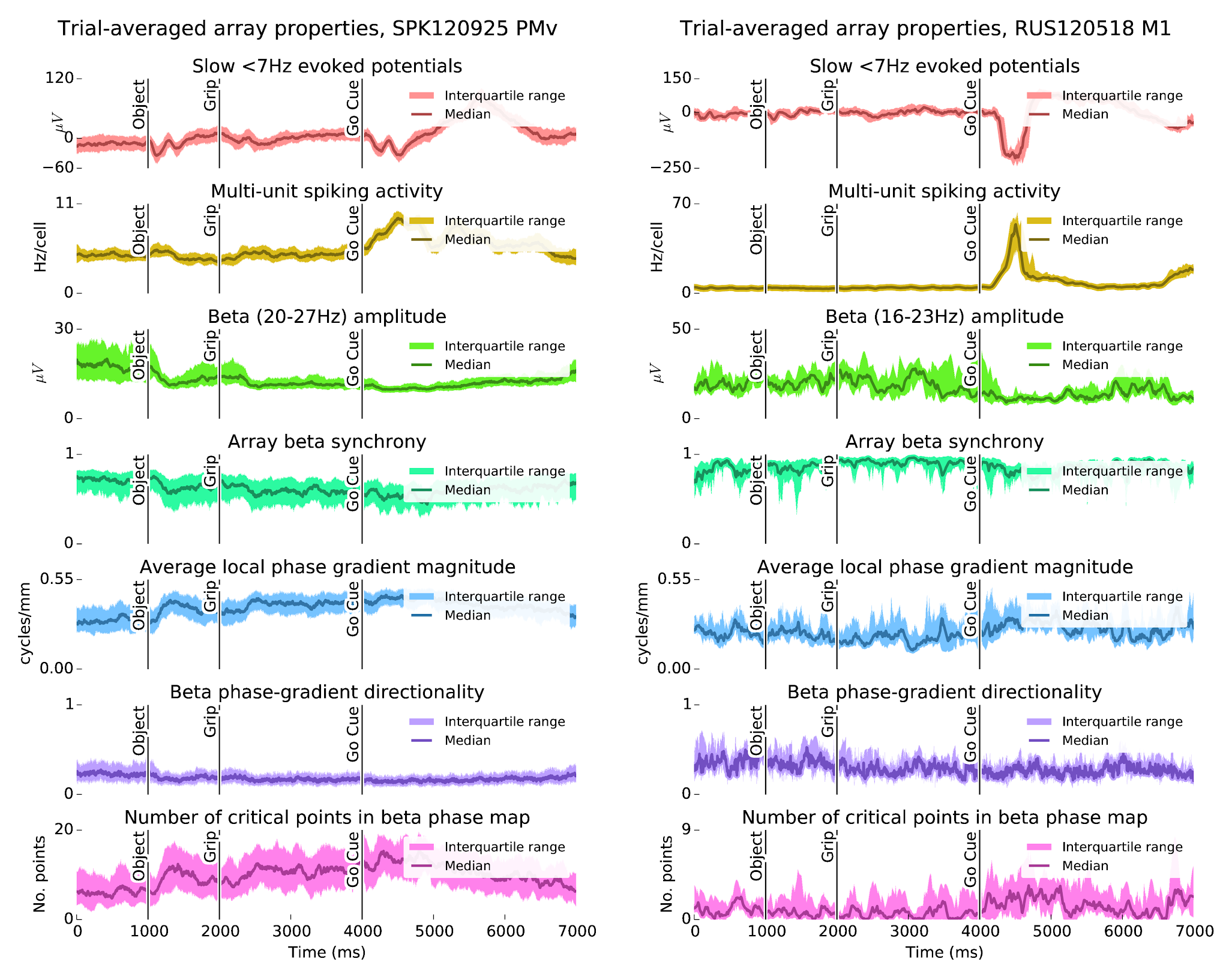
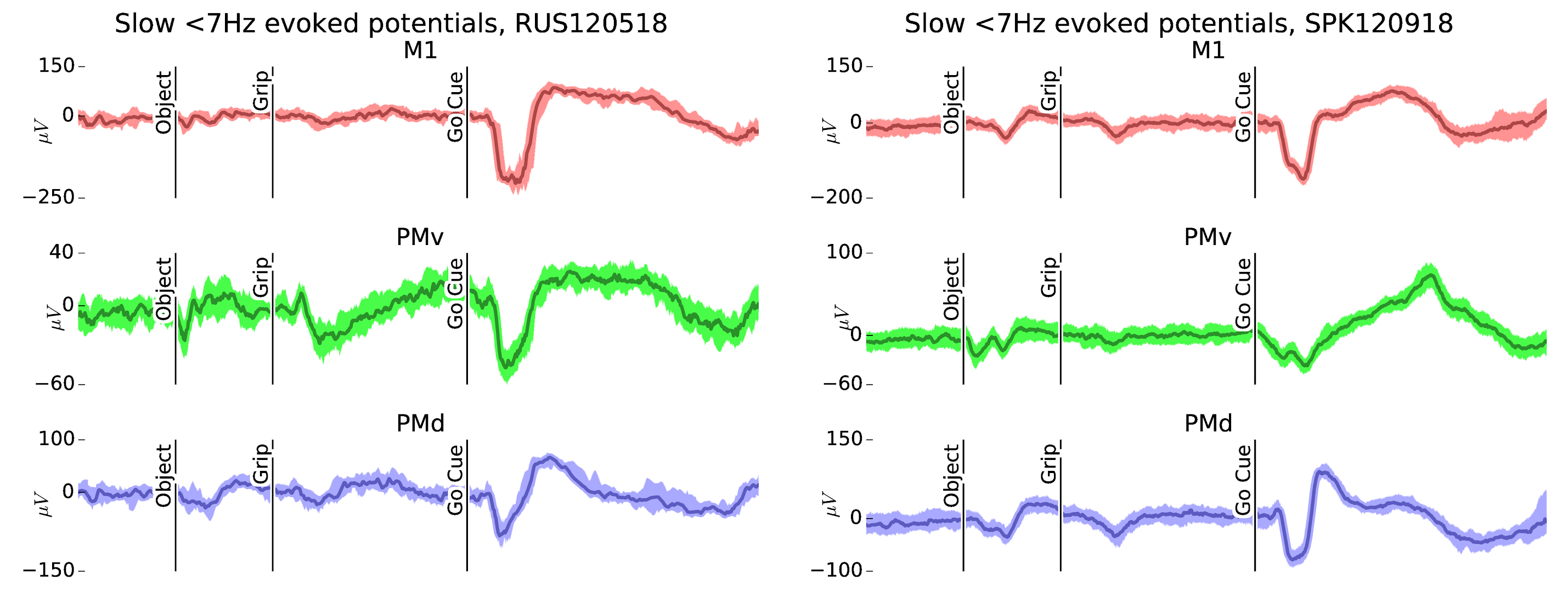
Figure 2: Evoked potentials in the CGID task. Both subjects show task-locked evoked potentials in the LFP (shown here averaged over good channels within a single area). In addition to strong, reliable motor evoked potentials in all areas, we also see visual-cue evoked potentials.
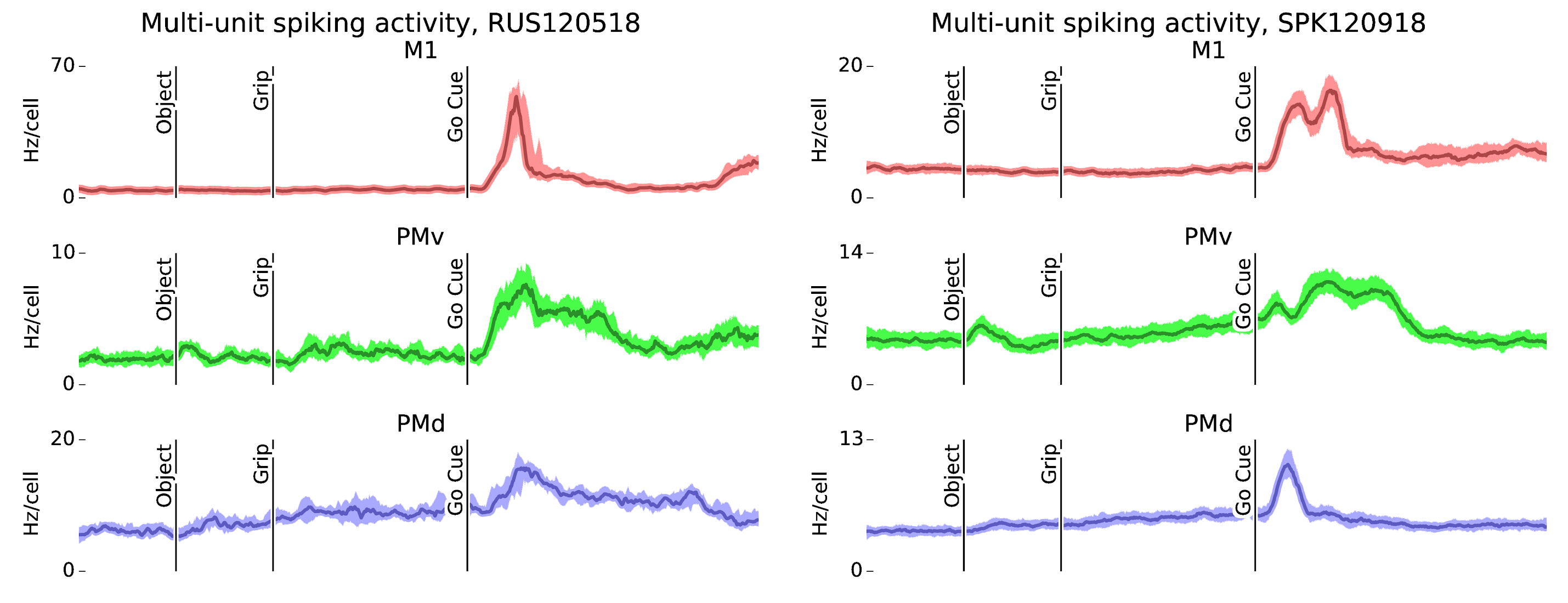
Figure 3: Task-locked modulations in multi-unit spiking activity vary between motor areas. In both subjects R and S, multi-unit spiking activity modulates to the CGID taks, for all motor areas. Only PMv displays multiunit modulation after object presentation. All motor areas display movement-evoked increases in firing rates, with area PMv showing elevated spiking extending for up to two seconds after go cue.
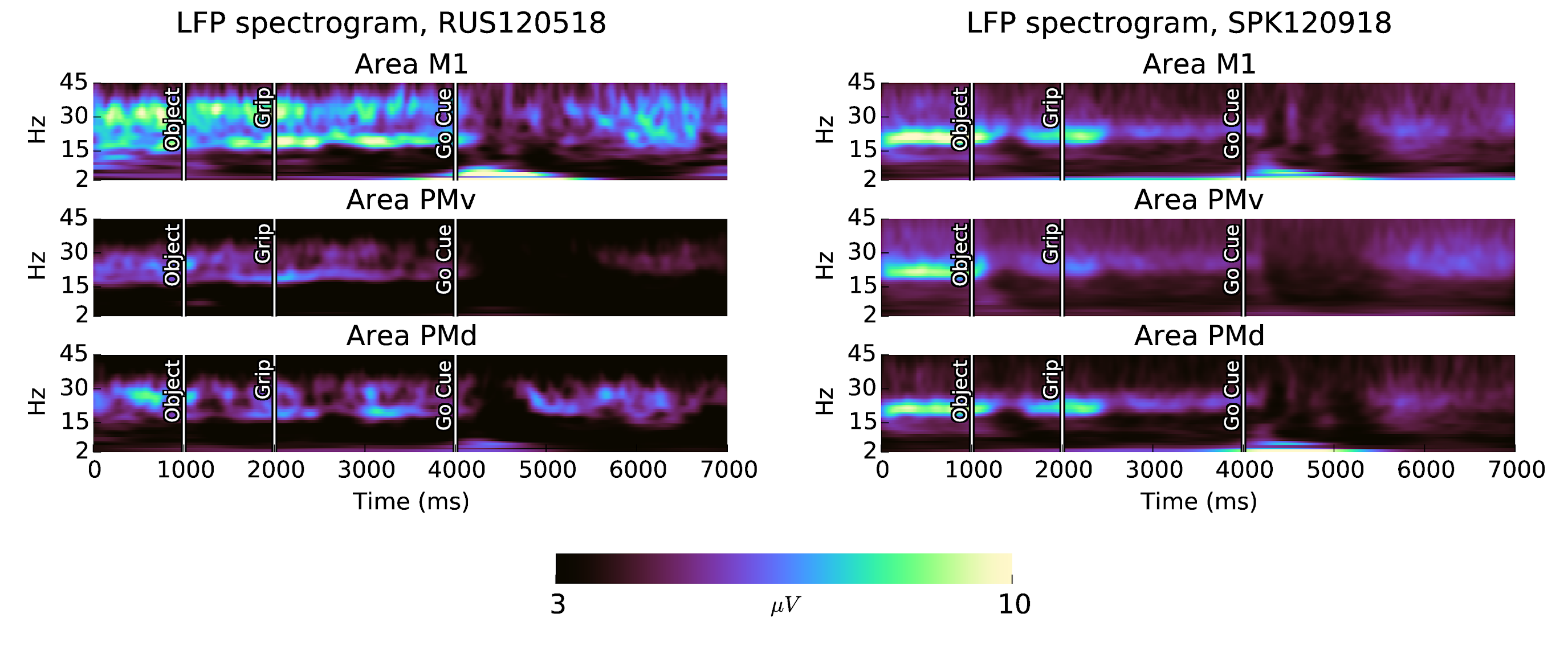
In subject S, beta oscillations are strongest before object presentation. In subject R, beta oscillations show greater power during the delay between the grip cue and the go cue (Fig. 4). There are some interesting features of the task-locked LFP activity:
- The frequency of beta oscillations increases during beta suppression in the planning-period in all areas of subject S, and in area PMd of subject R. In subject R, the frequency of beta oscillations after movement is also elevated.
- Visual cues evoke beta suppression and low frequency evoked potentials, much like movement.
- There is a short spike in alpha (7-15Hz) LFP activity immediately following object presentation in al areas with exception of subject R area PMd.
- Motor evoked potentials do not consist entirely of a slow <2Hz component, but also contain separate identifiable peaks in the theta (2-7Hz) band, in all areas except subject S area PMv.
- For subject S, the beta suppression evoked by object presentation is strongest in area PMv.
- The duration of movement-related beta suppression is longer in areas M1 and PMv than in area PMd, consistant with speculation that PMd is concerned with visually guided reaching, which completes before the object manipulation movements.
- The beta band in subject R is more complex, consisting of two bands which are enharmonic. The changes in the relative power in these two bands corresponds to changes in the time-domain appearance of the shape of the beta LFP waveform.
Figure 4: Beta oscillations are prominent in the CGID task. All areas in both subjects show increased beta LFP power during the instructed delays in the CGID task. The movement period after the go cue correlates with beta suppression and motor evoked potentials. Beta suppression also occurs after visual cue presentation to a lesser extent. Spectra are computed using a complex Morlet wavelet transform, with scale parameter of 10. Channels with excessive noise, and trials containing artefacts, are excluded.
Task-locked modulations in beta spatiotemporal activity
The spatiotemporal properties of beta oscillations vary throughout the CGID task. Beta synchrony is elevated during the planning periods, but is especially strong surrounding the visual cues, and absent during periods of beta suppression (Fig. 5). Although beta oscillations in subject R are composed of two enharmonic frequency bands, it is the lower 15-25 Hz fundamental frequency that displays spatial synchrony.
Figure 5: In the CGID task, LFP spatial synchrony occurs in the alpha, beta, and low-frequency bands. Beta oscillations are typically more synchronous over each area at times when beta power is high. Periods of beta power suppression correlate with reduction in beta synchrony. In both subjects a brief period of alpha-band (5-10Hz) synchrony appears at the start of beta suppression, most prominently in area PMv after object presentation. Synchrony in lower frequency (<7Hz) LFP is present throughout the entire task, both during the planning and movement periods. Because area PMv in subject S is much less synchronous than others, and area PMd in subject S much more synchronous, these plots have been z-scored so that the variations in synchrony over time and frequency can be compared across areas.

Figure 1 details the task-locked changes in the spatiotemporal structure of beta oscillations in area PMv for one session from subject S. This figure focus on the narrow 20-27 Hz band containing the beta LFP power for this session. Beta suppression approximately 250 ms after each of the visual cues is apparent, as is movement related beta suppression. Beta oscillations are attenuated during the laset 750 ms of the planning period, though not as much as during movement. Spatial synchrony shows more trial-to-trial variability and relatively less task-locking than beta amplitude. Similarly, weaker task locking is evident in other measures of beta spatiotemporal structure, such as the average magnitude of spatial gradient of Hilbert phase, the phase gradient directionality measure from Rubino et al. (2009), and the number of critical points in the Hilbert phase map.
Figure 6: Beta synchrony is not locked to the CGID task. Shown here are phase locking values (0 = no phase locking, 1 = complete phase locking) over all sessions and channels. We see phase-locking of synchronous alpha oscillations following the object presentation and go cue, as well as reproducible phase in motor evoked potentials.
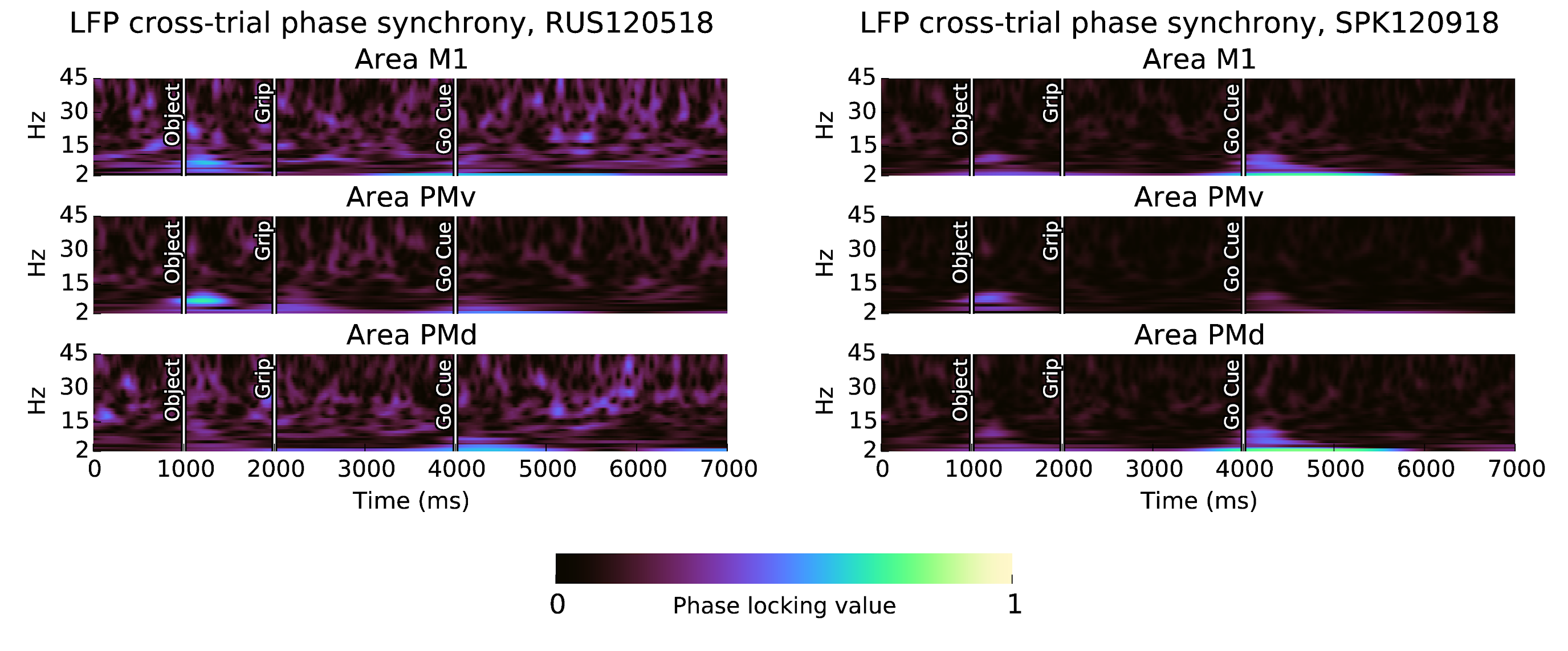
Overall, in subject S area PMv, beta oscillations before object presentation are synchronous, consisting primarily of global synchrony, and increases in phase gradient directionality accompanied with reduction in the average local phase gradient, indicative of traveling plane waves. Object presentation is followed by a suppression in beta amplitude, and correlates with a reduction in spatial synchrony, and an increase in the complexity of the beta spatiotemporal structure, based on increases in the average local phase gradient, and in the number of critical points in the Hilbert phase map. Beta LFP amplitude increases again surrounding the grip cue, accompanied by a modest increase in synchronization and reduction in the complexity of beta spatiotemporal patterns. Beta oscillations are suppressed again approximately 300 ms after grip cue, and remain reduced during the remainder of the planning period. The movement period is accompanied by a nearly complete suppression of beta amplitude, as well as a dramatic increase in spiking activity and a large motor-evoked potential. Beta spatiotemporal activity remains asynchronous during the movement period.
Correlations between beta spatiotemporal statistics and other measures of collective activity in motor cortex
During the instructed delays, we elevated beta power in motor cortex is associated with greater spatial synchrony. Both visual cue presentation and movement correlate with beta suppression, as well as increased single unit firing, slow <7 Hz LFP potentials, reduced beta synchrony, and increased in the complexity of spatiotemporal wave structure in the beta band (as measured by the phase gradient directionality and the number of critical points in the Hilbert phase map).
However, beta LFP activity is variable from trial to trial. Beta oscillations typically occur in transient events, lasting of 1-3 cycles. The precise nature and timing of beta spatiotemporal patterns is seldom reproduced across trials. Patterns consist of travelling and complex waves that evolve dynamically. To assess the spatiotemporal structure of beta frequency LFP oscillations, we correlate various measures of spatiotemporal activity and population dynamics, across trials over 50 ms time-bins.
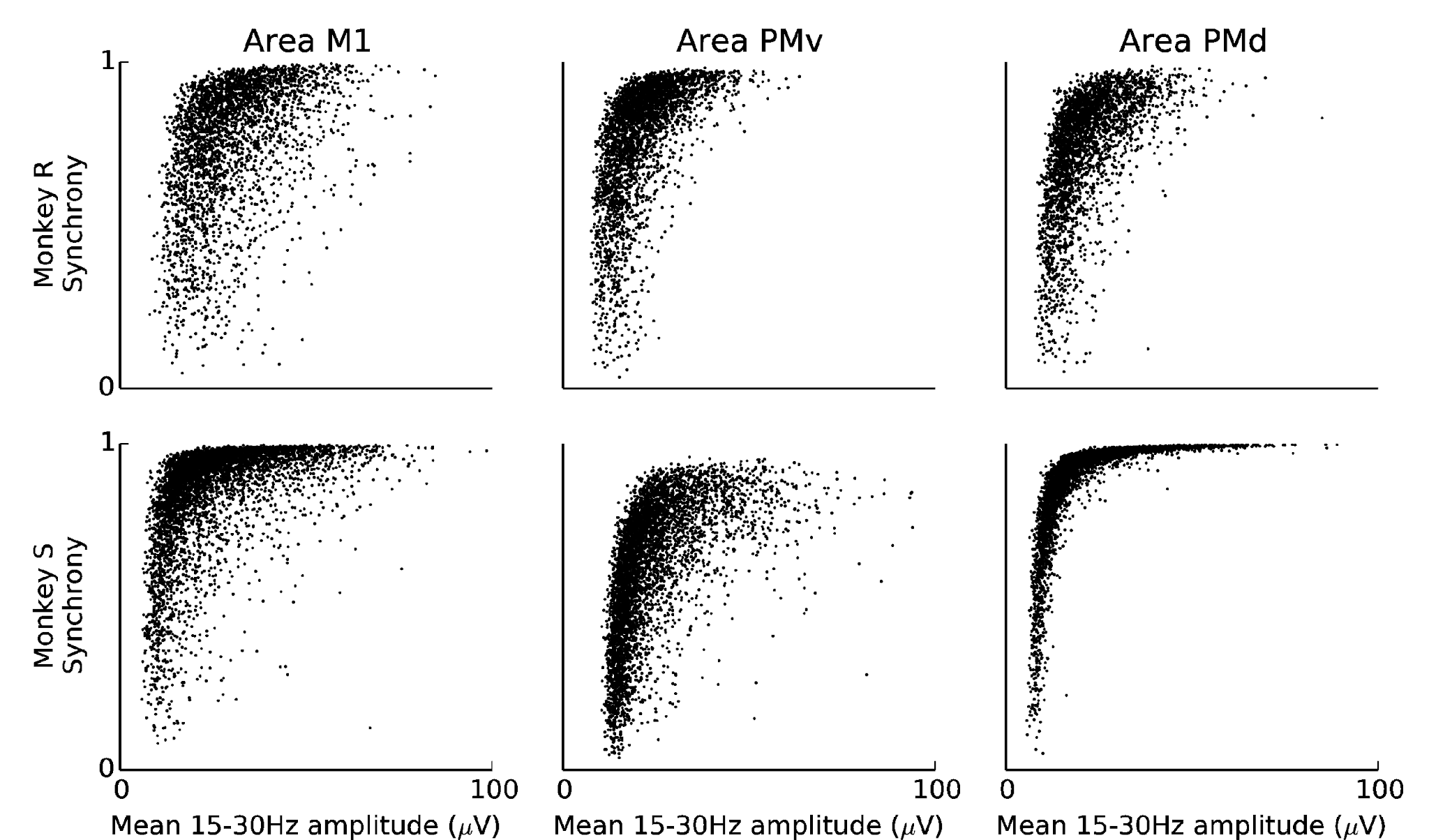
For example, Figure 7 demonstrates that periods of high beta amplitude tend to be highly synchronous, although this is not uniformly the case, with many high-amplitude time periods appearing asynchronous, and for example area PMv in subject S showing less synchrony than area PMd.
Beta synchrony correlates with high beta amplitude. In these plots, all sessions are combined for each subject and area. Synchrony and amplitude are averaged over 50ms time-bins.
Overall,
Beta oscillations progress from asynchrony to uniform synchrony via emergence of plane wave dynamics that approach spatially homogeneous beta phase
We find that beta oscillations transition from low-amplitude asynchronous states, to high-amplitude synchronous states, in the following manner:
locally asynchronous activity with becomes ordered, and phase singularities are eliminated,
correlation length scales and measure of wavelength increase with increasing beta amplitude,
the system progresses to a spatially synchronized state through gradual lengthening of wavelength.
No comments:
Post a Comment